This article was first published in 1988 in an issue of Social Research based on a conference we organized at The New School entitled, “In Time of Plague: The History and Social Consequence of Lethal, Epidemic Disease,” held at the height of the hysteria surrounding the outbreak of HIV-AIDS. The issue attempted to put that epidemic in perspective by considering it in the context of the social history of past lethal epidemics. Unfortunately, times are such that republishing this issue seemed appropriate. It will appear with a set of new comments about Covid-19 as the summer issue of Social Research.
Pasteur and Darwin
Charles Darwin’s role in nineteenth-century thought, how that shapes our own thinking about man’s place in Nature, is too well-known and oft-discussed to bear extensive elaboration on my part. His contemporary, Louis Pasteur, is a culture hero, world-renowned for the human benefits of his germ theory of disease: the use of antiseptic hygiene and of vaccines to prevent infection.
The ideological interaction of these two iconoclasts has been given too little attention. In his correspondence, Darwin makes enthusiastic but passing reference to Pasteur’s humanitarian contributions. Pasteur’s correspondence has been less extensively indexed to date. The most notable allusion to Darwin in his published work is his address to the Sorbonne on April 7, 1864: “Great problems are in question today, which keep all spirits in suspense: … the creation of man several thousand years or several thousand centuries ago; the fixity of species, … the idea of a useless God.” There is little doubt he is referring to Charles Darwin, whose work had been translated into French in 1862 and promptly aroused a theological storm. Pasteur is determined, however, to remove himself from that debate and such mysteries. Instead, he insists on addressing only those questions accessible to experiment, namely, the contemporary claims of spontaneous generation of microbial life. In 1864, his refutation was in comfortable support of an orthodoxy that would invoke the Creator for the ultimate origin of life. Indirectly, it was an argument against a Darwinian evolution of life arising in “some warm little pond.” Pasteur did show that the plethora of empirical claims of abiogenesis in sterilized broths exposed to air could all be accounted for by airborne spread of existing germs. By 1883, he had returned more optimistically to mechanistic views of abiogenesis if only one could achieve biochemical asymmetries, perhaps by the use of electromagnetism. Nevertheless, there is no record that he ever achieved a sympathetic understanding of Darwinian evolutionary theory; and he seems always to have been hostile to a methodology of inference, like Darwin’s, that deviated from the grain of laboratory experiment.
On Darwin’s side, for all his appreciation of Pasteur’s medical contributions, he seems never to have incorporated microbiology into his natural history. And, as we know, neither of them had any inkling of two other contemporaries’ contribution to fundamental biological understanding. Gregor Mendel’s foundations of genetics, articulated in 1865, were buried until 1900. Friedrich Miescher had discovered nucleic acids (DNA) in pus cells in 1870; we were not to begin to understand the biological function of DNA until Avery, MacLeod, and McCarty’s work at the Rockefeller Institute in 1944. The latency of DNA research may be ascribed mainly to deficiencies in experimental technique whose repair needed decades of drudgery and many instrumental inventions. The barriers between Darwin, Pasteur, and Mendel were purely cerebral and ideological.
What lost opportunity! Darwin might have found, as present-day investigators do, marvelous experimental material for the study of evolution in populations of microbes—where generation time is measured in minutes, and where natural (or artificial) selection can be applied to tens or hundreds of billions of unicellular organisms at small cost and less ethical compunction. Pasteur and his successors in microbiology might have avoided decades of muddled thinking about variation in bacteria. The revolution in biotechnology could have had a couple of decades’ head start. I should not complain: I had the fun and advantage in 1946 of exploring a terra still incognita (genetics of bacteria) that might otherwise have been blanketed with homestead claims for four or five prior decades.
Plagues
Darwin had placed Homo sapiens at the pinnacle of the evolutionary process, but with as much emphasis on pinnacle as on evolution. He never quite rectified the view that man has a privileged place in nature. Man’s intelligence, his culture, his technology has, of course, left all other plant and animal species out of the competition. Darwin was oblivious about microbes as our competitors of last resort. In experimental science, the Darwinian and Pasteurian perspectives are at last fully integrated. The study of mechanisms of virulence is a top priority in research laboratories applying the most advanced techniques of molecular genetics. Since Theobald Smith in 1934, F. M. Burnet and R. Duboshave offered us broad perspectives of the natural history of infectious disease-perspectives that leave no illusions about the feasibility of eradicating our scourges, of the ongoing struggle. For a period, the works of Paul de Kruif dramatized the efforts of the “microbe-hunters.” But one legacy of the “miracle drugs,” the antibodies of the 1940s, has been an extraordinary complacency on the part of the broader culture. Most people today are grossly overoptimistic with respect to the means we have available to forfend global epidemics comparable to the Black Death of the fourteenth century (or, on a lesser scale, the influenza of 1918), which took a toll of millions of lives! We have no guarantee that the natural evolutionary competition of viruses with the human species will always find ourselves the winner.
I would ask the professional cultural historians for their comment; but it appears that our half-century has turned away from external nature and to the self-deprecation of human nature, or of human organizations, as the central target of fear and struggle. Not that we have to quarrel over pride of place between virus infection and nuclear doomsday.
The countercultural protest against technology posits a benign nature, whose balance we now disturb with diabolical modernities. But man himself is a fairly recent emergent on the planet; the sheer growth of our species since the paleolithic is the major source of disturbances to that hypothetical balance. Man as a creature of culture is a man-made species; for better or worse, the only planet we know is a Promethean artifact. Genesis mandates: “Be fruitful and multiply!” After sampling the tree of knowledge, and acquiring the means, we could return to Eden only by reducing the human population to about 1 percent of its current density. We are complacent to trust that nature is benign; we are arrogant to assert that we have the means to except ourselves from the competition. But our principal competitors for dominion, outside our own species, are the microbes: the viruses, bacteria, and parasites. They remain an interminable threat to our survival.
This harsh view may be a product of my day-to-day laboratory experience. Most of my own scientific contributions have entailed the relentless use of artificial selection as a way to detect rare differences in the genetic makeup of individuals in large populations. These were populations of bacteria, but they numbered in the billions in each test tube. Typically, all but a few of these would be wiped out by the chemical or virus intentionally added to remove that “normal background”; a few survivors of uncharacteristic genetic composition are then readily detected and isolated. So, I have personally observed, even contrived, the wipeout of populations on a gigascale, and of course recognize that this is an unremitting process in nature—for example, recovery from infection on the part of any patient. This may come about either by the administration of an antibiotic or the mobilization of the naturally evolved defense mechanisms of the patient. In such confrontations, either the human individual or billions of microbes must die.
Is there any reason to believe that such processes are unique to the test tube, that life in the large is exempt from them? Of course not! Only the time scale is certain to be different, by a factor of years to minutes, of a million to one, the disparity of generation time of human to bacteria. The fundamental biological principles are the same. The numerical odds may be different, by a factor hard to estimate.
As crowded as we are, humans are more dispersed over the planetary surface than are the “bugs” in a glass tube, and we have somewhat fewer opportunities to infect one another, jet airplanes notwithstanding. The culture medium in the test tube offers fewer chemical and physical barriers to virus transmission than the space between people—but you will understand why so many diseases are sexually transmitted. The ozone shield still lets through enough solar ultraviolet light to make aerosol transmission less hospitable, and most viruses are fairly vulnerable to desiccation in dry air. The unbroken skin is an excellent barrier to infection; the mucous membranes of the respiratory tract much less so. And we have evolved immune defenses, a wonderfully intricate machinery for producing a panoply of antibodies, each specifically attuned to the chemical makeup of a particular invading parasite. In the normal, immune-competent individual, each incipient infection is a mortal race: between the penetration and proliferation of the virus within the body, and the development of antibodies that will dampen or extinguish the infection. If we have been vaccinated or infected before with a virus related to the current infection, we can mobilize an early immune response. But this, in turn, provides selective pressure on the virus populations, encouraging the emergence of antigenic variants. We see this most dramatically in the influenza pandemics, and every few years we need to disseminate fresh vaccines to cope with the current generation of the flu virus.
Many quantitative mitigations of the pandemic viral threat are then inherent in our evolved biological capabilities of coping with these competitors. Mitigation is also built into the evolution of the virus: it is a pyrrhic victory for a virus to eradicate its host! This may have happened historically, but then both that vanquished host and the victorious parasite will have disappeared. Even the death of the single infected individual is relatively disadvantageous, in the long run, to the virus-compared to a sustained infection leaving a carrier-free to spread the virus to as many contacts as possible. From the virus’s perspective, its ideal would be a virtually symptomless infection, in which the host is quite oblivious of providing shelter and nourishment for the indefinite propagation of the virus’s genes. Our own genome probably carries hundreds of thousands of such stowaways. The boundary between them and the “normal genome” is quite blurred; intrinsic to our own ancestry and nature are not only Adam and Eve but any number of invisible germs that have crept into our chromosomes. Some confer incidental and mutual benefit. Others of these symbiotic viruses (or “plasmids”) have reemerged as oncogenes, with the potential of mutating to a state that we recognize as the dysregulated cell growth of a cancer. As much as 95 percent of our DNA may be “selfish,” parasitic in origin.
At evolutionary equilibrium, we would continue to share the planet with our parasites, paying some tribute but deriving some protection from them against more violent aggression. Such an equilibrium is unlikely on terms we would voluntarily welcome: at the margin, the comfort and precariousness of life would be evenly shared. No theory lets us calculate the details; we can hardly be sure that such an equilibrium for earth even includes the human species. Many prophets have foreseen the contrary, given our propensity for technological sophistication harnessed to intraspecies competition.
In fact, innumerable perturbations remind us that we cannot rely on “equilibrium”—each individual death of an infected person is a counterexample. Our defense mechanisms do not always work; viruses are not always as benign as would be predicted to serve their long-term advantage.
The historic plagues, the Black Death of the fourteenth century, the recurrences of cholera, the 1918 swine influenza should be constant reminders of nature’s sword over our head. They have been very much on my mind for the past two decades. However, when I have voiced such fears, they have been mollified by the expectation that modern hygiene and medicine would contain any such outbreaks. There is, of course, much merit in those expectations: the plague bacillus is susceptible to antibiotics, and we understand its transmission by rat-borne fleas. Cholera can be treated fairly successfully with simple regimens like oral rehydration (salted water with a touch of sugar). Influenza in 1918 was undoubtedly complicated by bacterial infections that could now be treated with antibiotics; and if we can mobilize them in time, vaccines can help prevent the global spread of a new flu. On the other hand, the role of secondary bacterial infection in 1918 may well be overstated: it is entirely possible that the virus itself was extraordinarily lethal. The retrospective scoffing at the federal campaign against the swine flu of 1976 is a cheap shot on the part of critics who have no burden of responsibility for a wrong guess. It underrates health officials’ legitimate anxiety that we might have been seeing a recurrence of 1918—and underscores the political difficulty of undertaking the measures that might be needed in the face of a truly species-threatening pandemic. This so-called fiasco, in fact, mitigated an epidemic that happily proved to be of a less lethal virus strain. The few cases of side-effects attributed to the (polyvalent) vaccine are undoubtedly less than would have appeared from the flu infections avoided by the vaccination program. However, the incentives to attach fault for damages from a positive intervention have predictable consequences in litigation, not to be confused with the balance of social costs and benefits of the program as a whole.
Many outbreaks of viral or bacterial infections have destroyed large herds of animals, of various species, usually leaving a few immune survivors. With all the discussion of faunal extinctions, nothing has been said about infectious disease. It would be impossible to verify this from the fossil record, but disease is the most plausible mechanism of episodic shifts in populations. Incontrovertible examples of species wipeouts are seen with fungi in the plant world: Dutch elm disease and the American chestnut blight. Yes, it can happen.
My discussion has emphasized viruses because medical science has still to develop effective drugs for the treatment of virus infections—we have but a small handful, of limited use. Keep in mind that bacteria are free-living organisms whose metabolic peculiarities lend themselves to differential attacks. For example, the bacterial cell wall is utterly unlike any structure found in human cells. Hence, penicillin, which attacks the integrity of the bacterial cell wall, is all but innocuous to human tissue and can be given in very large doses so as to saturate every susceptible bacterial cell. Viruses, on the other hand, are genetic fragments that live within the host cells and exploit their metabolism. It has so far been very difficult to find chemicals that will inhibit a virus without harming the host cell at the same time. Our principal strategy for dealing with viruses is immunization, evoking antibodies that recognize the peculiarities of the virus surface. When a virus, like AIDS, comes along and targets the immune system itself, we are left with dimmer hopes of being able to use that strategy; and we have very few alternatives.
AIDS and Other Plagues
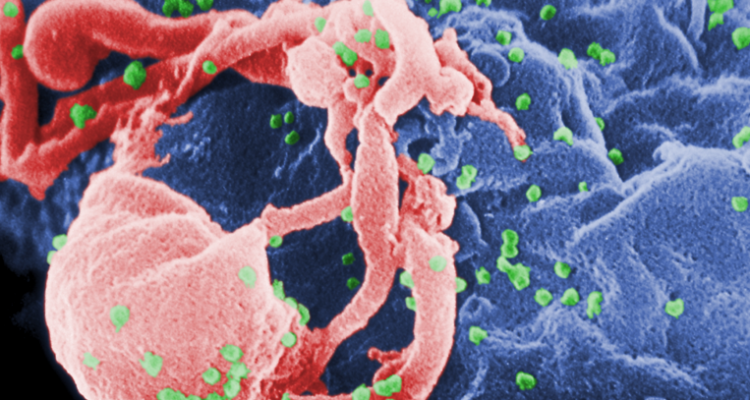
The sudden and tragic spread of AIDS has brought us back to earth in our speculation on plagues: who among us has not been personally touched already! A host of social and ethical issues come right to home and will be the main focus of this conference. As always, the third world is paying the heaviest price, in the dying of whole villages and in the stigma of the biological origins of the virus. We are all in fear of what will come next. Will the virus spread still further? What are the prospects of a vaccine? Of a cure?
We are all too familiar with the factors that have made AIDS an especially ugly challenge. Unlike other virus infections, which leave some survivors immune to further attack, there is nothing in the natural history of AIDS to point either to a cure or to a vaccine. Victims develop antibodies, then go on notwithstanding to develop more aggravated disease, with the eventual collapse of the immune system. The fact that this is still mysterious makes it the most promising avenue for new discovery and possible intervention. Most of the factual knowledge we have is unremittingly discouraging.
The long latent period multiplies the opportunity for spread; the victim may be unaware of carrying the virus, even less his contacts. Nothing could provoke more anxiety than this protracted uncertainty. The targeting of the immune system also encourages the seeding of other infections—we are already starting to see a recrudescence of tuberculosis in the United States and aggravations of syphilis and a host of opportunistic organisms rarely seen before. It would be far worse were HIV still more readily spread, but its substantial confinement to special high-risk groups worsens the social tensions around efforts at control. The long latent period guarantees a large number of momentarily healthy carriers whose civil rights—for example, to continued schooling and employment—are in instant conflict with a quarantine mentality for public-health control. I was labeled an alarmist twenty years ago for raising a “specter” of a pandemic. My most pessimistic imagination did not fetch the constellation of attributes that we observe with AIDS. AIDS is already so prevalent in the United States today that none of the approaches of public-health control of other acute infections are pertinent. There is little merit in targeting a handful of individuals, generally the most compliant, when there is a score as many freely walking the streets.
So much is unknown about AIDS that a large amount of testing is essential just to understand the scope and localization of the problem. We may soon find that many hospital and medical procedures aggravate AIDS infection: that will obligate broader testing of AIDS among hospital admissions, simply for the patients’ benefit. And they will sue for not having been routinely tested. Health-care personnel have an ethical obligation to care for all the sick but this is complemented by a right to know what they need to protect their own health. We will not work out the most viable balance between individual rights and the community’s needs without a great deal more compassionate thought and inevitable political stress. Both need to be informed by more reliable knowledge.
Our preoccupation with AIDS should not obscure the multiplicity of infectious diseases that threaten our future. It is none too soon to start a systematic watch for other new viruses before they become so irrevocably lodged. The fundamental bases of virus research can hardly be given too much encouragement—and they have made extraordinary leaps, particularly with the help of recombinant DNA technology. Such research should be done on a broad international scale, both to share the progress made in advanced countries and to amplify the opportunities for fieldwork in the most afflicted ones.
We also have some political lessons learned. Hard-won human rights, the autonomy of individuals, will be in conflict with the quietude of the community. At severe cost, if at all, will it be possible to impose traditional disease-control methods like isolation and quarantine on new viruses. Compulsory vaccination has all but passed the pale. Claims for redress for individual harm from medical accidents from vaccines necessitate that we find new social-insurance approaches to indemnification. Failing that, we have already seen a collapse of the pharmaceutical industry’s incentive and capability for pursuing new vaccine developments. The most stunning victories will be quiet ones, against viruses we have learned enough about beforehand to keep them from planting a foothold.
The stresses on democratic civility posed by AIDS have no precedent in U.S. history. They are compounded by our scientific uncertainties as to where this epidemic is heading. The best available advice is incorporated in the program advocated by the leadership of the federal health agencies and the expertise of groups like the National Academy of Sciences. That advice can be no more authentic than the empirical findings to date. It is of the greatest urgency that these be bolstered by a more robust appreciation of virus biology and of the human immune mechanism. At present, nothing we know gives us assurance of finding satisfactory cures or vaccines for AIDS infection. We can take small comfort that much more remains to be explored—but only if we mount that exploration with the most urgent priority.
Joshua Lederberg, 1925-2008, was a microbiologist. Awarded a Nobel Prize in 1958, he served as president of Rockefeller University from 1978-1990.
This condensed essay was originally published in Social Research, Volume 55, No. 3 (Fall 1988), pp. 343–59, where it can be found with citations and in its complete form. It is reprinted here by permission.